

C57BL/6JNifdc-Fcgr2btm1(FCGR2B)Bcgen/Bcgen • 112052
| Product name | B-hCD32B mice |
|---|---|
| Catalog number | 112052 |
| Strain name | C57BL/6JNifdc-Fcgr2btm1(FCGR2B)Bcgen/Bcgen |
| Strain background | C57BL/6JNifdc |
| Aliases | CD32, CD32B, FCGR2, FCG2 |
Gene targeting strategy for B-hFCD32B mice. The exons 2-3 of mouse Fcg2b gene that encode the extracellular domain were replaced by human FCGR2B exons 3-5 in B-hCD32B mice.
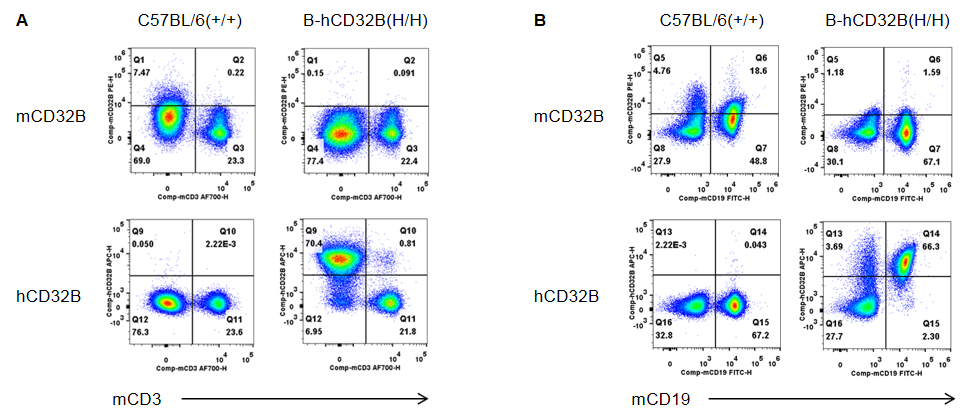
Strain specific CD32B expression analysis in wild-type mice and homozygous B-hCD32B mice by flow cytometry. Blood cells were collected from wild-type C57BL/6 mice and homozygous B-hCD32B mice. (A) Mouse CD32B was not detectable in T cells of wild-type mice. Human CD32B was not detectable in T cells of homozygous B-hCD32B mice. (B) Mouse CD32B was detectable in B cells of wild-type mice. Human CD32B was detectable in B cells of homozygous B-hCD32B mice.
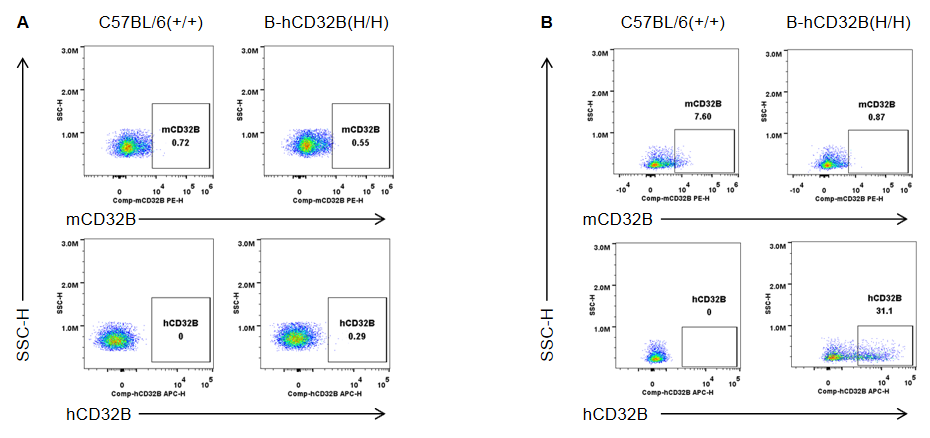
Strain specific CD32B expression analysis in wild-type mice and homozygous B-hCD32B mice by flow cytometry. Blood cells were collected from wild-type C57BL/6 mice and homozygous B-hCD32B mice. (A) Mouse CD32B was not detectable in granulocytes of wild-type mice. Human CD32B was not detectable in granulocytes of homozygous B-hCD32B mice. (B) Mouse CD32B was detectable in monocytes of wild-type mice. Human CD32B was detectable in monocytes of homozygous B-hCD32B mice.
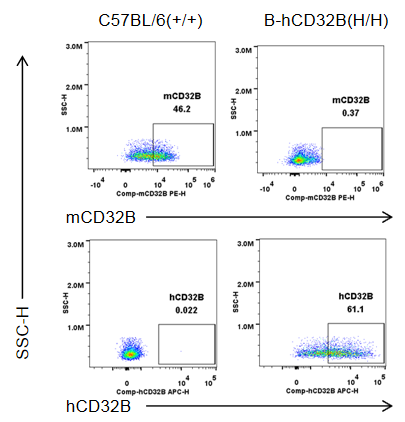
Strain specific CD32B expression analysis in wild-type mice and homozygous B-hCD32B mice by flow cytometry. Blood cells were collected from wild-type C57BL/6 mice and homozygous B-hCD32B mice. Mouse CD32B was detectable in macrophages of wild-type mice. Human CD32B was detectable in macrophages of homozygous B-hCD32B mice.
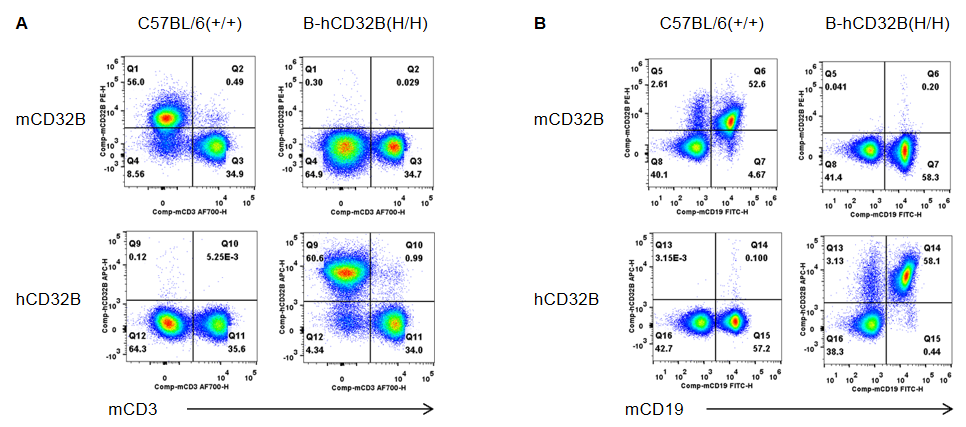
Strain specific CD32B expression analysis in wild-type mice and homozygous B-hCD32B mice by flow cytometry. Splenocytes were collected from wild-type C57BL/6 mice and homozygous B-hCD32B mice. (A) Mouse CD32B was not detectable in T cells of wild-type mice. Human CD32B was not detectable in T cells of homozygous B-hCD32B mice. (B) Mouse CD32B was detectable in B cells of wild-type mice. Human CD32B was detectable in B cells of homozygous B-hCD32B mice.
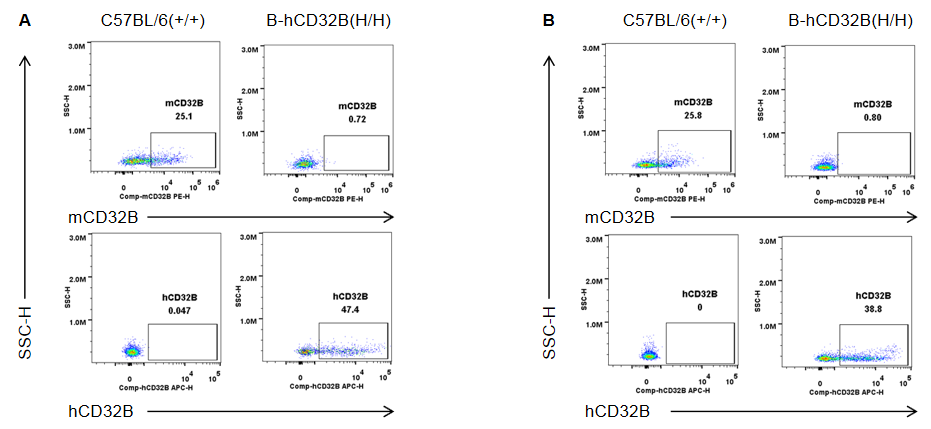
Strain specific CD32B expression analysis in wild-type mice and homozygous B-hCD32B mice by flow cytometry. Splenocytes were collected from wild-type C57BL/6 mice and homozygous B-hCD32B mice. (A) Mouse CD32B was detectable in DC of wild-type mice. Human CD32B was detectable in DC of homozygous B-hCD32B mice. (B) Mouse CD32B was detectable in monocytes of wild-type mice and heterozygous B-hCD32B mice. Human CD32B was detectable in monocytes of heterozygous B-hCD32B mice.
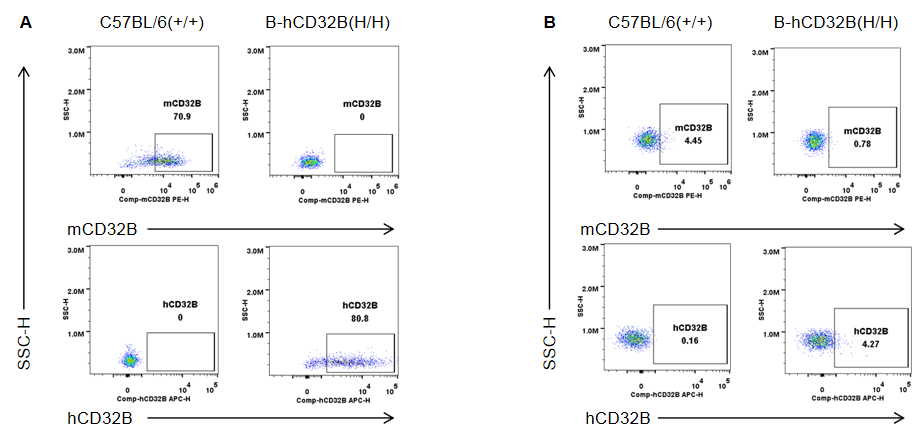
Strain specific CD32B expression analysis in wild-type mice and homozygous B-hCD32B mice by flow cytometry. Splenocytes were collected from wild-type C57BL/6 mice and homozygous B-hCD32B mice. (A) Mouse CD32B was detectable in macrophages of wild-type mice. Human CD32B was detectable in macrophages of homozygous B-hCD32B mice. (B) Mouse CD32B was weakly detectable in granulocytes of wild-type mice. Human CD32B was weakly detectable in granulocytes of homozygous B-hCD32B mice.
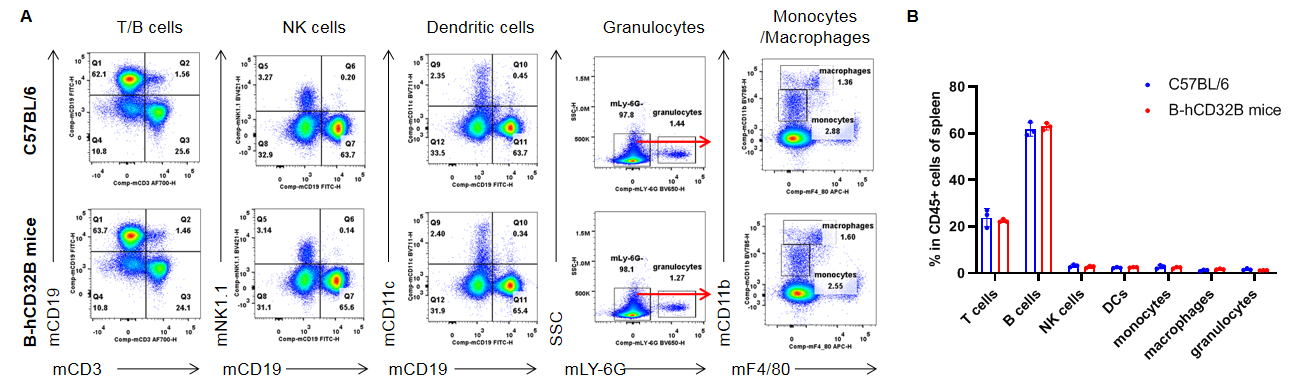
Analysis of the spleen leukocyte subpopulations by FACS. Splenocytes were collected from WT and homozygous B-hCD32B mice (n=3, 7-week-old). Flow cytometry analysis of the splenocytes was performed to assess leukocyte subpopulations. A. Representative FACS plots. Single live cells were gated for the CD45+ population and used for further analysis as indicated. B. Results of FACS analysis. The percentages of T cells, B cells, NK cells, dendritic cells, granulocytes, monocytes, and macrophages in homozygous B-hCD32B mice were similar to those in the C57BL/6 mice, demonstrating that introduction of hCD32B in place of its murine counterpart does not change the overall development, differentiation or distribution of these cell types in the spleen. Values are expressed as mean ± SEM.
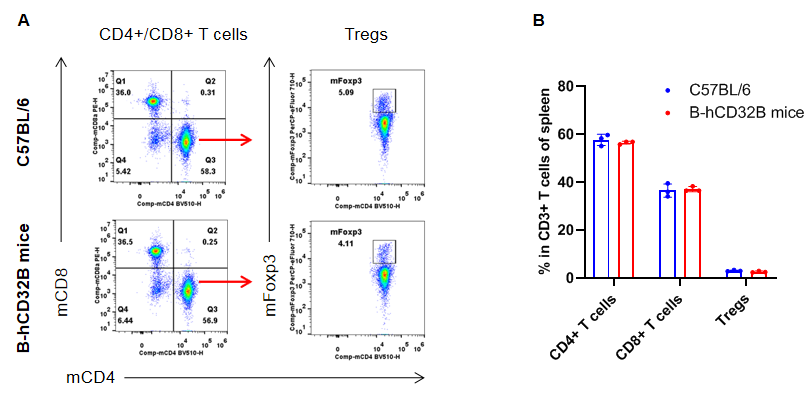
Analysis of the spleen T cell subpopulations by FACS. Splenocytes were isolated from female C57BL/6 and homozygous B-hCD32B mice (n=3, 7-week-old). Flow cytometry analysis of the splenocytes was performed to assess leukocyte subpopulations. A. Representative FACS plots. Single live CD45+ cells were gated for CD3+ T cell population and used for further analysis as indicated. B. Results of FACS analysis. The percentages of CD8+ T cells, CD4+ T cells, and Tregs in homozygous B-hCD32B mice were similar to those in the C57BL/6 mice, demonstrating that introduction of hCD32B in place of its murine counterpart does not change the overall development, differentiation or distribution of these T cell subtypes in the spleen. Values are expressed as mean ± SEM
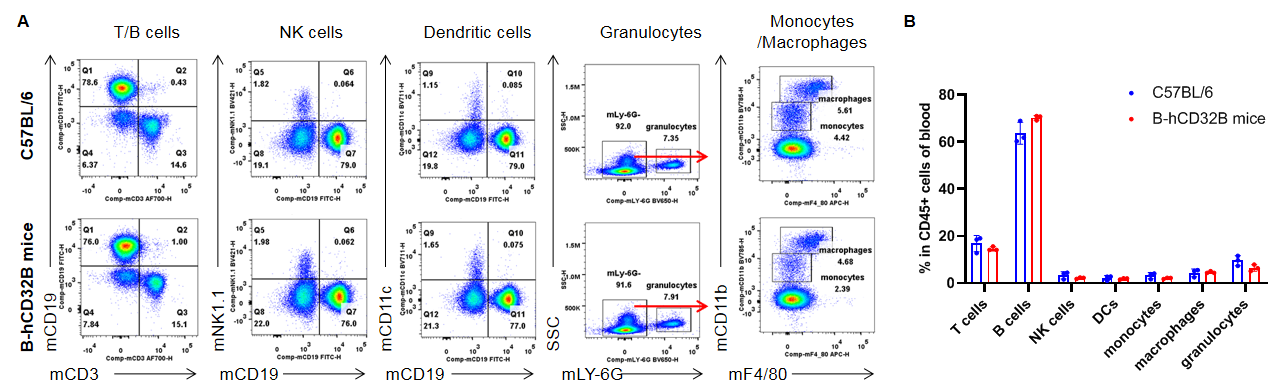
Analysis of the blood leukocyte subpopulations by FACS. Blood cells were collected from WT and homozygous B-hCD32B mice (n=3, 7-week-old). Flow cytometry analysis of the blood was performed to assess leukocyte subpopulations. A. Representative FACS plots. Single live cells were gated for the CD45+ population and used for further analysis as indicated. B. Results of FACS analysis. The percentage of T cells, B cells, NK cells, dendritic cells, granulocytes, monocytes, and macrophages in homozygous B-hCD32B mice were similar to those in the C57BL/6 mice, demonstrating that introduction of hCD32B in place of its murine counterpart does not change the overall development, differentiation or distribution of these cell types in the blood. Values are expressed as mean ± SEM.
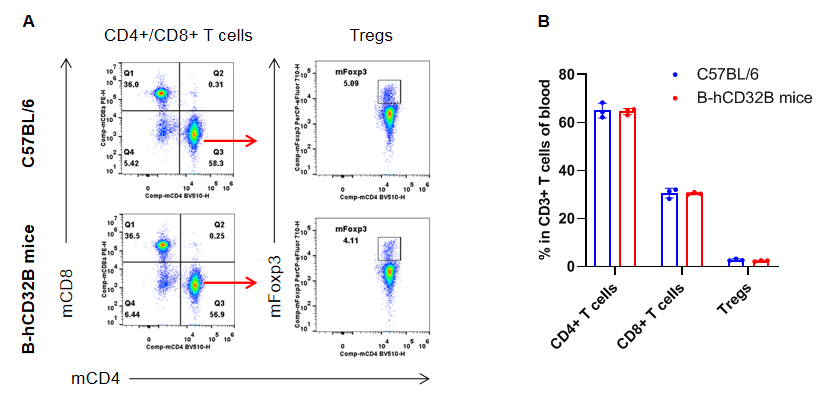
Analysis of the blood T cell subpopulations by FACS. Blood cells were isolated from female C57BL/6 and homozygous B-hCD32B mice (n=3, 7-week-old). Flow cytometry analysis of the blood was performed to assess leukocyte subpopulations. A. Representative FACS plots. Single live CD45+ cells were gated for CD3+ T cell population and used for further analysis as indicated. B. Results of FACS analysis. The percentages of CD8+ T cells, CD4+ T cells, and Tregs in homozygous B-hCD32B mice were similar to those in the C57BL/6 mice, demonstrating that introduction of hCD32B in place of its murine counterpart does not change the overall development, differentiation or distribution of these T cell subtypes in the blood. Values are expressed as mean ± SEM.
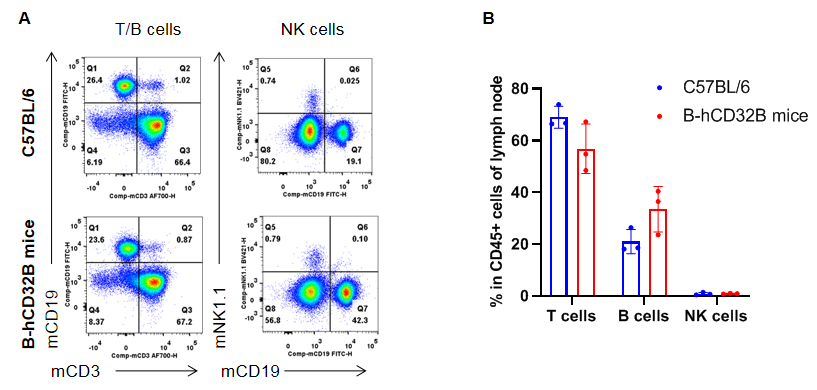
Analysis of lymph nodes leukocyte subpopulations by FACS. Lymph nodes were collected from WT and homozygous B-hCD32B mice (n=3, 7-week-old). Flow cytometry analysis of the lymph nodes was performed to assess leukocyte subpopulations. A. Representative FACS plots. Single live cells were gated for the CD45+ population and used for further analysis as indicated. B. Results of FACS analysis. The percentages of T cells, B cells, and NK cells in homozygous B-hCD32B mice were similar to those in the C57BL/6 mice, demonstrating that introduction of hCD32B in place of its murine counterpart does not change the overall development, differentiation or distribution of these cell types in the lymph nodes. Values are expressed as mean ± SEM.
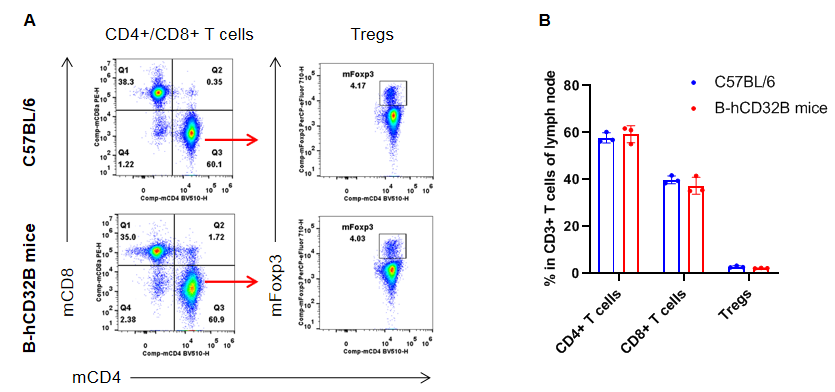
Analysis of lymph nodes T cell subpopulations by FACS. Lymph nodes were isolated from female C57BL/6 and homozygous B-hCD32B mice (n=3, 7-week-old). Flow cytometry analysis of the lymph nodes was performed to assess leukocyte subpopulations. A. Representative FACS plots. Single live CD45+ cells were gated for CD3+ T cell population and used for further analysis as indicated. B. Results of FACS analysis. The percentages of CD8+ T cells, CD4+ T cells, and Tregs in homozygous B-hCD32B mice were similar to those in the C57BL/6 mice, demonstrating that introduction of hCD32B in place of its murine counterpart does not change the overall development, differentiation or distribution of these T cell subtypes in the lymph nodes. Values are expressed as mean ± SEM.
